【转载】贵州大学发现CMV劫持宿主蛋白形成“秘密基地”,并找到精准打击的小分子
2026-06-03 浏览次数: 10
2026年,Molecular Plant Pathology发表了贵州大学赵磊、宋润江等团队的研究成果。他们发现CMV衣壳蛋白(CP)与宿主钙依赖性蛋白激酶CDPK7-like互作,通过液-液相分离形成凝聚体,促进病毒增殖。CDPK7-like过表达增强CMV侵染,却抑制PVY和PMMoV,显示病毒特异性“劫持”。进一步鉴定出香豆素衍生物D3靶向CP的Thr52残基,破坏CP-CD PK7-like互作及凝聚体形成,抗CMV活性优于利巴韦林(EC₅₀=70.8 μg/mL),且对蜜蜂低毒、植物安全。该研究揭示了病毒利用相分离操控宿主的新机制,并提供新型抗病毒先导化合物。
研究流程全景图:
Co-IP-MS筛选CMV CP互作蛋白 → 转录组联合分析锁定CDPK7-like → LCA/BiFC验证互作并发现凝聚体 → FRAP/1,6-己二醇证实LLPS性质 → 基因沉默/过表达/转基因验证CDPK7-like促CMV侵染 → 合成香豆素衍生物D1-D34 → 抗病毒活性筛选(D3最优)→ DARTS靶标鉴定→分子对接/MD模拟确认Thr52关键位点 → 点突变(T52A)验证Thr52对侵染和D3活性的必要性

一、为什么是突破?——“病毒-宿主相分离”成为抗病毒新靶点
植物病毒与宿主蛋白形成相分离凝聚体促进侵染的报道罕见。本研究首次揭示:CMV CP与CDPK7-like互作形成LLPS液滴,该过程依赖CP的Thr52残基;CDPK7-like过表达特异增强CMV侵染(病毒RNA增3.16倍),却抑制PVY和PMMoV,显示CMV“劫持”该激酶。更重要的是,香豆素衍生物D3靶向Thr52,拆散凝聚体,EC₅₀仅70.8 μg/mL,优于利巴韦林(195 μg/mL)。这为靶向病毒-宿主相分离的抗病毒农药开发提供了全新策略。
二、环环相扣的设计逻辑——从互作筛选到小分子干预
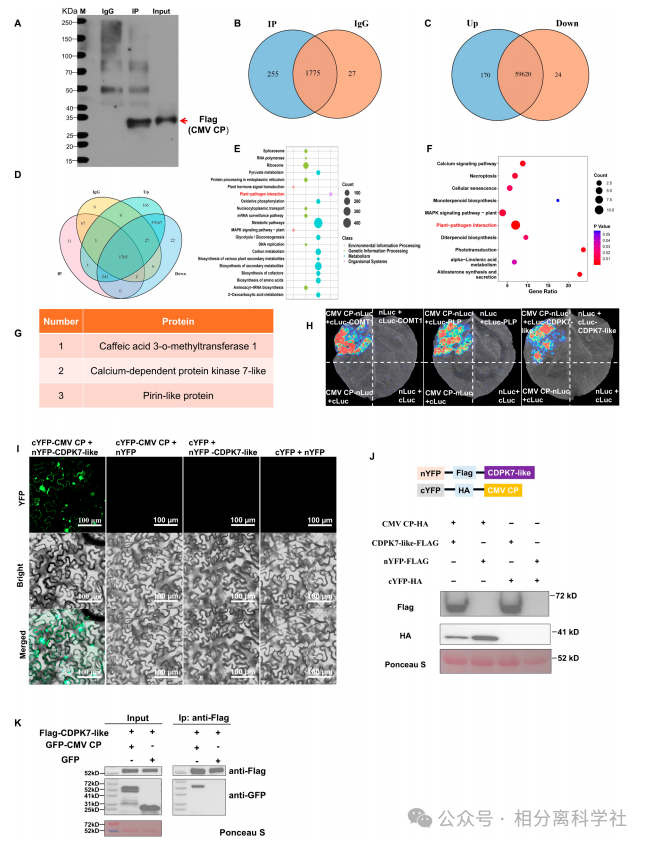
第一步:筛选CMV CP宿主互作蛋白
Co-IP-MS鉴定出255个候选互作蛋白,KEGG富集于“植物-病原互作”“钙信号通路”。转录组分析发现CP表达上调170个基因、下调24个。两者交集获3个候选:COMT1、PLP、CDPK7-like。→ 锁定CDPK7-like为潜在免疫相关因子。
第二步:验证互作与相分离
LCA和BiFC证实CMV CP与CDPK7-like直接互作,且仅此组合产生大量凝聚体。FRAP显示荧光恢复(τ~数十秒),1,6-己二醇处理消除凝聚体,证明LLPS性质。凝聚体定位于核膜和质膜。→ 首次报道CMV CP-宿主激酶相分离。
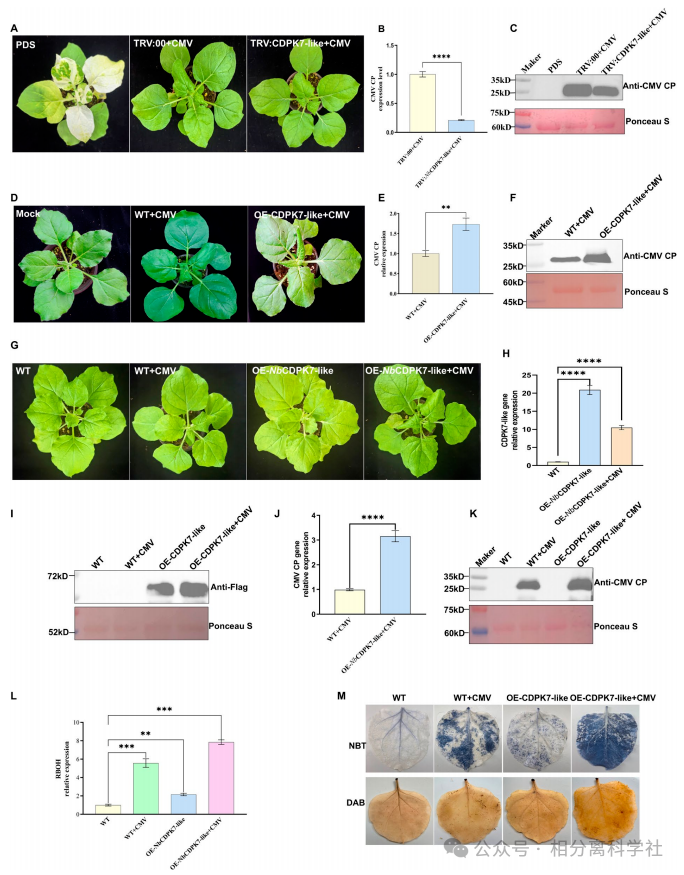
第三步:功能验证——CDPK7-like促CMV侵染
基因沉默CDPK7-like:CMV CP RNA降8.06倍,症状减轻。瞬时/稳定过表达:CMV RNA增1.7-3.16倍,症状加重。但过表达抑制PVY(RNA降0.46倍)和PMMoV(0.41倍),且PVY/PMMoV CP不与CDPK7-like互作。→ 证明CMV特异性劫持该激酶,且与相分离相关。
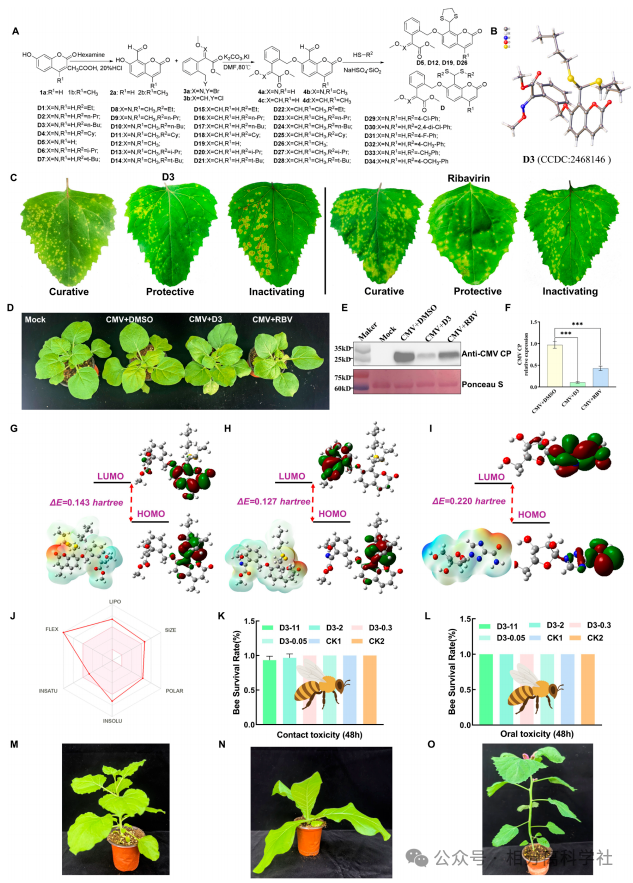
第四步:设计小分子D3靶向Thr52
合成34个香豆素衍生物,D3抗CMV活性最优(EC₅₀=70.8 μg/mL)。DARTS实验证实D3保护CMV CP免受蛋白酶降解。分子对接和MD模拟显示D3与CP的Thr52形成关键氢键;T52A突变后结合亲和力下降(Kd从9.3 nM升至20.1 nM),且T52A突变病毒在植株中症状显著减轻、RNA积累降低。D3对T52A突变病毒无效。→ Thr52是CMV侵染关键位点,也是D3作用靶点。
三、核心验证——从相分离机制到小分子体内药效
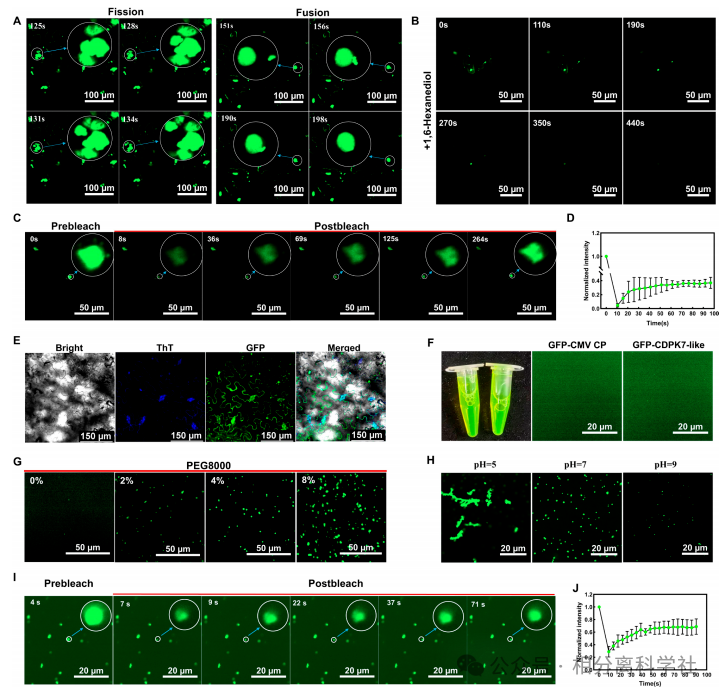
相分离的体内证据
BiFC中CMV CP-CD PK7-like共表达产生大量动态液滴,FRAP显示荧光恢复(证明液态),1,6-己二醇处理后液滴消失。体外纯化蛋白同样形成液滴并发生融合。→ 确认LLPS驱动凝聚体形成。
CDPK7-like的双向调控
过表达CDPK7-like增强CMV却抑制PVY/PMMoV,且只有CMV CP与之互作。这暗示CMV进化出“劫持”该激酶的策略,而其他病毒可能被该激酶抑制。凝聚体的形成可能屏蔽了CDPK7-like对防御信号的正常传导。
小分子靶标确证
DARTS显示D3专一保护CMV CP(其他蛋白无保护条带)。分子对接显示D3与Thr52形成氢键,MD模拟中T52A突变导致配体RMSD波动增大、结合不稳。T52A突变病毒在烟草原生症状几乎消失(仅轻微褪绿),且D3对其无效。→ 确证Thr52是D3的关键作用残基。
农用化学品的成药性
D3对蜜蜂急性经口/接触LD₅₀>11 μg/蜂(EPA无毒标准),500 μg/mL喷施本氏烟、烟草、苋色藜无药害。SwissADME预测其类药性良好。→ 具备农药开发潜力。
四、方法学优势——为什么这套“发现-验证-干预”策略值得借鉴?
多组学联用:Co-IP-MS+转录组交集,缩小候选范围,避免假阳性。
BiFC意外发现:BiFC不仅验证互作,还揭示凝聚体现象,提示LLPS可能性。
FRAP+1,6-己二醇+体外重构:三重证据确证相分离。
宿主功能双向验证:沉默+过表达+转基因,且对比其他病毒,证明特异性劫持。
小分子靶标鉴定:DARTS+分子对接+点突变+MD模拟,形成靶标确证闭环。
毒理学快速评估:蜜蜂毒性+植物安全性,筛选早期排除高风险化合物。
五、意义与展望
该研究首次揭示CMV通过CP与宿主CDPK7-like相分离形成凝聚体促进侵染的机制,并发现靶向CP关键残基Thr52的小分子D3,为抗植物病毒农药研发提供了全新靶点和先导结构。相分离作为病毒-宿主互作的新维度,未来或成为抗病毒药物设计的通用平台。
未来方向:
解析CP-CD PK7-like凝聚体的高分辨率结构,指导更精准的药物设计。
优化D3结构,提高田间药效和稳定性。
探索其他植物病毒是否也存在类似的相分离劫持机制。
评估D3与现有抗病毒剂复配的增效潜力。
一句话总结: CMV用“相分离液滴”锁住宿主激酶为自己服务,小分子D3拆散这个“保护罩”,让植物重获抵抗力。